- Что нужно знать об эксцентрических упражнениях
- Эксцентрический тренинг как инструмент для наращивания мощи и мышечной массы
- Эксцентрический тренинг – что это
- Достоинства тренинга
- Оптимальное время растягивания эксцентрической фазы
- Оптимальный вес эксцентрического опускания
- Включение эксцентрических нагрузок в программу тренировок
- Сравнение эксцентрического и концентрического тренинга
- Повреждение мышц после эксцентрических упражнений
- Проске У., Морган Д.Л.
- ПОВРЕЖДЕНИЕ МЫШЕЦ ПОСЛЕ ЭКСЦЕНТРИЧЕСКИХ УПРАЖНЕНИЙ:МЕХАНИЗМ, МЕХАНИЧЕСКИЕ КРИТЕРИИ, АДАПТАЦИЯ И КЛИНИЧЕСКИЕ ПРИЛОЖЕНИЯ
- РЕЗЮМЕ
- НАЧАЛЬНЫЕ СОБЫТИЯ
- СТРУКТУРНЫЕ ПРИЗНАКИ
- СДВИГ ОПТИМУМА ДЛИНЫ САРКОМЕРА
- СНИЖЕНИЕ НАПРЯЖЕНИЯ
- РАЗЛИЧНЫЕ ТИПЫ МЫШЕЧНЫХ ВОЛОКОН
- ПОВЫШЕНИЕ ПАССИВНОГО НАПРЯЖЕНИЯ
- НАБУХАНИЕ МЫШЦЫ И БОЛЕЗНЕННОСТЬ
- АДАПТАЦИЯ
- ОРГАНЫ КИНЕСТЕЗИИ
- КЛИНИЧЕСКИЕ ПРИМЕНЕНИЯ
- ЗАКЛЮЧИТЕЛЬНЫЕ КОММЕНТАРИИ
Что нужно знать об эксцентрических упражнениях
В этой статье мы собрали предыдущие посты Татьяны Зайченко об эксцентрических упражнениях.
Определение
Эксцентрическим называют сокращение, при котором напряжённая мышца удлиняется под нагрузкой.
Это происходит в уступающей фазе движения, когда мы вынуждены уступать силе гравитации и/или весу снаряда.
В большинстве случаев эксцентрическое сокращение реализуется при движении вниз (приседания, выпады, наклоны, подтягивания/тяги в наклоне, отжимания/жимы — то есть любая работа со свободными весами и собственным весом, где решает гравитация). Исключение составляют упражнения с использованием резинок или блочных тренажеров — там эксцентрическая работа идет в направлении тяги резинки или рукоятки тренажера, например, вверх (тяга сверху) или вперед, как при тяге к поясу сидя.
При эксцентрическом сокращении мышечно-сухожильный комплекс растягивается и поглощает механическую энергию.
Далее возможны два сценария:
1) После растягивания не происходит последующего укорочения мышцы, и энергия рассеивается в виде тепла.
Пример: спрыгивание с турника. При приземлении мышцы растянулись, сдерживая чрезмерное сгибание сегментов тела, т.е. эксцентрически сократились, погасив ударную нагрузку. Занавес.
2) После растягивания мышца укорачивается (то есть происходит переход от эксцентрического сокращения к концентрическому), что позволяет использовать эластическую энергию растяжения для увеличения силы концентрического сокращения. Чем быстрее этот переход, тем выше эффективность сокращения. Быстрый переход от эксцентрического сокращения к концентрическому называется циклом растяжения-сокращения.
Пример: серия прыжков в отскок. При приземлении мышцы голеней растянулись, сдерживая чрезмерное тыльное сгибание голеностопа, и я тут же оттолкнулась стопами от пола (мышцы голени немедленно укоротились, сгибая стопы в подошвенном направлении) и снова подпрыгнула вверх.
ЭУ широко используются:
- в период реабилитации после травм — разумеется, по назначению компетентного специалиста
- после вынужденных перерывов в тренировках, не связанных с травмой (в качестве «втягивающей» нагрузки)
- по ходу тренировочного процесса (в циклах анатомической адаптации)
Преимущества эксцентрических упражнений
1) Мышечная гипертрофия
При выполнении эксцентрических упражнений высокой интенсивности с продолжительностью сокращения 2-4 сек мышечная масса увеличивается за счёт роста поперечника волокон и числа саркомеров, преимущественно волокон II типа. Этот эффект более выражен у нетренированных людей.
2) Рост мышечной силы
Эксцентрические упражнения высокой интенсивности и низкого объёма обеспечивают значительное увеличение силы. Это актуально как для новичков, так и для тренированных спортсменов. Кроме того, использование ЭУ увеличивает взрывную силу, а результативность в прыжках повышается даже без выполнения специальных прыжковых упражнений.
3) Увеличение сократительной части мышцы без визуального увеличения длины
Это происходит за счёт добавления последовательно соединённых саркомеров в составе отдельных миофибрилл.
а) увеличивается скорость сокращения;
б) пик усилия мышца развивает раньше.
Это позволяет мышцам лучше предохранять суставы от повреждений при быстрых движениях – мышцы начинают сокращаться раньше, увеличивая контроль над движением. Крайне важная функция для профилактики травм и реабилитации.
4) Улучшение нервно-мышечной координации
В эксцентрической фазе мышечные волокна сокращаются в ответ на внешнее раздражение, то есть мышца «подстраивается» под навязанный ей сценарий. Таким образом, нервная система вынуждена обучаться быстрее, но вместе с тем увеличивается риск перенапряжения ЦНС.
Выводы
Включение эксцентрических упражнений в программу тренировок оптимизирует увеличение мощности, силы и массы мышц, длину мышц при развитии усилия, а также координацию движений.
Эксцентрические мышечные сокращения необходимы для поглощения ударной нагрузки во время приземления после прыжка, а также для увеличения эффективности прыжков, бросков, экономичности бега и ходьбы.
Внешняя нагрузка, которая вынуждает мышцу сокращаться эксцентрически, может превышать силу максимального концентрического сокращения на 20-50%. То есть, вес, с которым вы способны опуститься в присед, всегда больше веса, с которым вы можете встать.
Повреждения мышц происходит в основном именно при эксцентрических сокращениях, но тут все зависит от дозировки. ЭУ широко применяются в реабилитации, например, при повреждении ахиллова сухожилия или для профилактики травм мышц задней поверхности бедра.
Источник
Эксцентрический тренинг как инструмент для наращивания мощи и мышечной массы
Популярность эксцентрического тренинга начала возрастать в начале 21 века. До конца прошлого тысячелетия такой вид тренировок практиковался гораздо реже. Это связано с тем фактом, что при эксцентрических нагрузках спортсмены получают больше всего травм. На сегодняшний день эксцентрический тренинг используется не только как эффективный способ наращивания мышечной массы, но и в реабилитации или профилактике травм.
Изначально интерес к эксцентрическим нагрузкам был вызван в 70-х годах 20 века статьями Артура Джоунса, публиковавшимися в популярных спортивных журналах. Там Джоунс описывал тактики эксцентрических тренировок, впоследствии даже разработал специальные тренажеры.
Эксцентрический тренинг – что это
 Условно все упражнения делятся на два этапа – концентрический и эксцентрический. Оба этапа подразумевают сокращение мышц, только на концентрическом этапе происходит укорачивание мышцы, а на эксцентрическом удлинение.
Условно все упражнения делятся на два этапа – концентрический и эксцентрический. Оба этапа подразумевают сокращение мышц, только на концентрическом этапе происходит укорачивание мышцы, а на эксцентрическом удлинение.
Самым простым примером является упражнение со штангой – при поднятии штанги происходит концентрическое сокращение, при опускании – эксцентрическое.
Оба этапа призваны спровоцировать прирост мышечного объема, но эффект от эксцентрических нагрузок более продуктивный. Исследования показывают, что этому есть два объяснения:
- кислота фосфатидная – химическое соединение, вырабатываемое организмом при растяжении мышцы. Соединение активирует выработку мышечного протеина, это способствует набору мышечной массы;
- клетки-сателлиты – «дремлющие» клетки, находящиеся в наружной области мышц. В ходе эксцентрического упражнения ткани нередко повреждаются. Именно повреждения «пробуждают» сателлиты – клетки притягиваются в пораженную область и преобразовываются в мышечные волокна, становясь частью мускулатуры.
Достоинства тренинга
 Эксцентрические тренировки – действенный инструмент для наращивания мускулатуры, профилактики и избавления от последствий травм. Основные преимущества упражнений:
Эксцентрические тренировки – действенный инструмент для наращивания мускулатуры, профилактики и избавления от последствий травм. Основные преимущества упражнений:
- прирост мышечного объема. Для гипертрофии мышцам нужно сначала повреждаться. Во время интенсивной эксцентрической тренировки поврежденные участки восстанавливаются спутниковыми (сателлитовыми) клетками. Они отдают свое ядро, умножая концентрацию миозина и актина в саркомере. Таким образом, мышца растет в поперечнике;
- увеличение мускульной силы. В ходе эксцентрического тренинга увеличивается мощность, взрывная сила;
- улучшение гибкости. Регулярные эксцентрические нагрузки способны увеличивать диапазон движения сустава, а также длину мышцы;
- профилактика травм. Тренировки провоцируют мышцы адаптироваться к новым нагрузкам. Со временем мышцы достигают приемлемого растяжения, что снижает риск травмироваться;
- ускорение метаболизма. Эксцентрический тренинг разгоняет обмен веществ. Эта особенность тренинга будет полезна тем, кто хочет сбросить вес посредством силовых упражнений.
Эксцентричный тренинг также эффективен для атлетов, достигших тренировочного плато. Исследования доказали, что «пробуждение» клеток-сателлитов способствует скорейшему сдвигу гипертрофии мышц в сторону увеличения.
Оптимальное время растягивания эксцентрической фазы
 В ходе экспериментов с эксцентрическими тренировками многие спортсмены замедляли негативную фазу вплоть до 6 секунд. Но подобные затягивания не принесли ожидаемых плодов, чрезмерной гипертрофии добиться не удалось.
В ходе экспериментов с эксцентрическими тренировками многие спортсмены замедляли негативную фазу вплоть до 6 секунд. Но подобные затягивания не принесли ожидаемых плодов, чрезмерной гипертрофии добиться не удалось.
Ученые пришли к выводу – быстрые эксцентрические повторы гораздо эффективнее медленных и лучше провоцируют факторы роста – размножение спутниковых (сателлитовых) клеток и выработку мышечного протеина.
Объясняют это явление «поперечные мостики», образующиеся при соединении миозина с актином. Без этого соединения невозможно сокращение мышц. Большое количество «мостиков» защищает мышечную ткань от повреждений. При замедленном движении в эксцентрической фазе организм располагает большим временем для создания «мостиков». Снижение травмирования тканей приводит к сокращению выработки мышечного белка и уменьшению количества активных сателлитов.
Исходя из этих фактов, специалистами было установлено оптимальное время «растягивания» эксцентрической фазы – от 1 до 3 секунд.
Оптимальный вес эксцентрического опускания
 Проводился эксперимент, в котором участвовали пауэрлифтеры. Они выжимали одинаковый вес сначала с упором на концентрическую фазу, затем на эксцентрическую. При сравнении результатов тренировок оказалось, что разница в приросте мышечной массы стремится к нулю.
Проводился эксперимент, в котором участвовали пауэрлифтеры. Они выжимали одинаковый вес сначала с упором на концентрическую фазу, затем на эксцентрическую. При сравнении результатов тренировок оказалось, что разница в приросте мышечной массы стремится к нулю.
Это доказывает, что для достижения продуктивности эксцентрического тренинга необходимо поднимать вес, превышающий максимально возможный при концентрической тренировке. Человек способен опустить снаряд приблизительно на 70% тяжелее того, который может поднять.
Рекомендуемый тренерами вес при эксцентрическом тренинге составляет 100-170% от рабочего максимума.
Включение эксцентрических нагрузок в программу тренировок
 Фактически любое упражнение может стать эксцентрическим, нужно просто увеличить период растяжения мышц. Примеры упражнений:
Фактически любое упражнение может стать эксцентрическим, нужно просто увеличить период растяжения мышц. Примеры упражнений:
- отжимания – замедлить опускание тела, ускорить поднятие;
- подтягивания – в прыжке ухватиться за турник, потом медленно выпрямлять руки, держа вес собственного тела;
- приседания – замедлить опускание, ненадолго задержаться в приседе, затем быстро встать.
Для исключения травм мышц и суставов, концентрическую фазу необходимо выполнять быстро, но плавно.
Если же перед спортсменом стоит цель добиться увеличения мышечной массы и прироста абсолютной силы, то акцент делается на упражнениях с опусканием максимальных весов.
Не стоит сразу начинать со своего предельного веса – это может повлечь за собой серьезные травмы.
Специалисты рекомендуют начинать с развития навыка «контролируемого опускания». Проще говоря, сначала необходимо научиться опускать вес за то время, которое запланировали заранее.
После приобретения навыка подконтрольного опускания веса, можно начинать эксцентрические упражнения с использованием 70% от максимального веса. Постепенно увеличивать вес до эксцентрического предела.
Так как тренировки проходят с использованием сверхтяжелых нагрузок работать необходимо в паре. Например, в упражнении со штангой, партнер сначала помогает ее поднять, а затем контролирует ее опускание.
Интенсивные тренировки такого рода вызывают отложенные боли, которые способны продолжаться на протяжении нескольких дней. Поэтому до полной адаптации мышц к нагрузкам не рекомендуется повторять упражнения чаще одного раза в неделю.
Сравнение эксцентрического и концентрического тренинга
Проводилось множество исследований, сравнивающих эффект от концентрической и эксцентрической силовых тренировок. Каждое из них доказывает преимущества эксцентрического тренинга для ускорения наращивания мышечной массы и мощности.
К примеру, одно из исследований выявило, что через сутки после эксцентрической тренировки количество клеток-сателлитов возросло на 27%, в то время как после концентрической осталось неизменно. При этом сателлиты образуются исключительно в мышечных волокнах, способных к быстрому росту, то есть способствуют укоренной гипертрофии.
Эксцентрический тренинг эффективен для наращивания мышечной массы и абсолютной силы, способен адаптировать и защитить мышцы от серьезных травм. В то же время такие тренировки травмоопасны, их стоит делать исключительно подконтрольно, используя рекомендации тренера. При правильном подходе к тренировкам результат не заставит себя долго ждать.
Источник
Повреждение мышц после эксцентрических упражнений
В статье «Повреждение мышц после эксцентрических упражнений» рассмотрены: механизм, механические критерии, адаптация, клинические приложения, а также причины повреждения мышц. Показано, что эксцентрическом упражнении сокращающаяся мышца насильственно удлиняется, поэтому мышцы нетренированных людей на следующий день становятся жесткими и воспаленными из-за повреждения мышечных волокон.
U. Proske and D. L. Morgan Muscle damage from eccentric exercise: mechanism, mechanical signs, adaptation and clinical applications //Journal of Physiology, 2001.– V. 537.– N2, P.333–345.
Проске У., Морган Д.Л.
ПОВРЕЖДЕНИЕ МЫШЕЦ ПОСЛЕ ЭКСЦЕНТРИЧЕСКИХ УПРАЖНЕНИЙ:МЕХАНИЗМ, МЕХАНИЧЕСКИЕ КРИТЕРИИ, АДАПТАЦИЯ И КЛИНИЧЕСКИЕ ПРИЛОЖЕНИЯ
РЕЗЮМЕ
В эксцентрическом упражнении сокращающаяся мышца насильственно удлиняется; в концентрическом упражнении она сокращается. В то время как концентрические сокращения начинают движения, эксцентрические сокращения замедляют или останавливают их. Уникальная особенность эксцентрического упражнения – то, что мышцы нетренированных людей становятся жесткими и воспаленными на следующий день из-за повреждения мышечных волокон. Этот обзор рассматривает два возможных начальных события, ответственных за последующее повреждение: повреждение в системе сцепления-сокращения возбуждения и разрушение на уровне саркомеров. Другие изменения, которые наблюдаются при выполнении эксцентрических упражнений: падение активного напряжения, изменение в оптимальной длине при развитии активного напряжения и повышение пассивного напряжения, должны в итоге, описать разрушение саркомеров, как начального события при повреждении. Кроме повреждения мышечных волокон существуют свидетельства повреждения сенсорных органов мышцы и ее проприорецепции. Второй период упражнения, спустя неделю после первого, производит намного меньше повреждения. Это – результат процесса адаптации. Один из предложенных механизмов адаптации — увеличение количества саркомеров в мышечных волокнах, что приводит ко вторичному изменению в оптимальной длине мышцы при развитии активного напряжения. Способность мышцы, быстро адаптироваться после повреждения от эксцентрических упражнений увеличивает важность клинических приложений умеренного эксцентрических упражнений, чтобы защитить мышцу от большого количества более сильных повреждений.
Любая форма упражнения, если оно выполнено достаточно энергично, может стать болезненной. Но только одна форма упражнения, эксцентрического упражнения, если мы непривычны к этому, делает наши мышцы жесткими и воспаленными на следующий день. В течение эксцентрического упражнения возбужденная мышца насильственно удлиняется. Одним из примеров эксцентрического упражнения, с которым обычно сталкиваются, является ходьба по наклонной плоскости. Когда мы идем по наклонной плоскости, четырехглавая мышца бедра контролирует степень сгибания колена против силы гравитации, поэтому в каждом шаге мышца подвергается эксцентрическому сокращению. Немедленно после того, как выполнено упражнение, никакой боли не ощущается. Она возникает через несколько часов и достигает максимума приблизительно через 48 часов. Это, как считают, возникает из-за повреждения мышцы, произведенного эксцентрическим упражнением.
Интересная и важная особенность — процесс адаптации. Второе выполнение эксцентрического упражнения, спустя неделю после первого, не делает мышцы такими жесткими и воспаленными, как после первого выполнения. Способность мышцы быстро приспособиться к повреждению от эксцентрического упражнения, чтобы предотвратить дальнейшее повреждение, обеспечивает возможность множества клинических приложений.
Эксцентрические упражнения и механизм их действия обсуждались во множестве обзоров (Armstrong et al. 1991; McHugh et al. 1999; Morgan & Allen, 1999; Warren et al. 2001; Allen, 2001). Краткий обзор литературы указывает, что к этой теме проявляется растущий интерес.
Цель этого обзора состоит в том, чтобы сосредоточить внимание на некоторых индикаторах повреждения от непривычных эксцентрических упражнений и их возможных механизмах. Обсуждение будет рассмотрено, каким образом могли бы использоваться такие индикаторы, чтобы оценить степень защиты, доступной для человека в случае дальнейшего применения эксцентрических упражнений и как применить эти знания в клинике.
НАЧАЛЬНЫЕ СОБЫТИЯ
Установлено, что есть два видимых признака повреждения в мышце немедленно после того, как она была подвергнута ряду эксцентрических сокращений: наличие разрушенных саркомеров в миофибриллах и повреждение системы «возбуждения-сокращения» (E-C) в системе сцепления филаментов. Однако непонятно, что является первичным. Мы предполагаем (см. также Morgan & Allen, 1999), что процесс повреждения начинается со сверхрастяжения саркомеров (рис. 1). Альтернативная гипотеза заключается в том, что начальным событием является повреждение компонентов системы «возбуждения-сокращения» (E-C) в процессе сцепления. В недавнем обзоре, Warren et al. (2001) сравнил эти гипотезы и объявил, что отказ системы сцепления (E-C), возникающий после эксцентрического упражнения, вызывает снижение уровня напряжения мышцы на 75% и более.
Повреждение, замеченное в течение первых нескольких дней после упражнения, приписывается авторами физическому разрушению элементов в мышце, испытывающих напряжение. Таким образом, основное предположение заключается в том, что первичное повреждение возникает в системе сцепления E-C, и только маленькая часть повреждения происходит на уровне саркомеров. Этот вывод поддерживают наблюдения, что в мышце мышей падение напряжения после упражнения может быть возвращено кофеином (Warren et al. 1993; Balnave & Allen, 1995). В первом из этих исследований, уровень напряжения был восстановлен посредством 50 мм, кофеина, который способствует выделению Ca 2+ из саркоплазматического ретикулума и приводит к развитию контрактуры мышцы. Во втором исследовании 10 мм кофеина использовался для потенциации напряжения в мышечных волокнах в ответ на прямое электрическое возбуждение. На основе этого было сделано заключение, что изменение в системе сцепления E-C мышечных волокон мыши может быть основной причиной падения напряжения (силы) после эксцентрических сокращений (Allen, 2001).
Однако все еще оставляет открытым вопрос, что является первичным: отказ сцепления E-C или разрушение саркомеров. Существуют различные эффекты потенциации. В мышечных волокнах лягушки (Morgan et al. 1996), и в мышце жабы (Talbot, 1997; Allen, 2001), падение напряжения не могло быть возвращено потенциацией, посредством выделения ионов Ca 2+ .
Более подробно влияние эксцентрических сокращений на мышцы описано в моих книгах «Гипертрофия скелетных мышц человека» и «Биомеханика мышц«
Существует гипотеза процесса разрушения саркомера (рис. 1). Как известно, существует область, где напряжение саркомера падает (из-за особенностей зависимости «длина – напряжение саркомера», Gordon et al. 1966). Как предположил Morgan (1990), при активном растяжении основное изменение длины мышцы будет осуществляться за счет самых слабых саркомеров в миофибрилле или, строго говоря, самыми слабыми полу-саркомерами. На спускающейся ветви кривой «длина-напряжение», эти саркомеры станут прогрессивно еще более слабыми и когда они достигают критической точки, они удлиняются быстро, неудержимо, к точке, где нет никакого наложения миофиламентов, где напряжение в пассивных структурах уравновешивает активное напряжение в смежных саркомерах, которые все еще имеют наложение миофиламентов. Этот процесс повторяется многократно, с растяжением следующего самого слабого саркомера и так далее. Постулируется, что сверхрастяжение саркомеров распределено случайно в мышечном волокне. В конце растяжения, когда мышца расслабляется, миофиламенты в большинстве сверхрастянутых саркомеров, принимают такую длину, что они могут возобновить свою нормальную функцию. Некоторые не в состоянии это сделать и остаются разрушенными (Talbot & Morgan, 1996). Постулируется, что в течение повторных эксцентрических сокращений число разрушенных саркомеров растет, пока не будет повреждена мембрана. Именно в этот момент повреждение элементов сцепления E-C, становится очевидным. Впоследствии волокно может умереть (рис. 1).
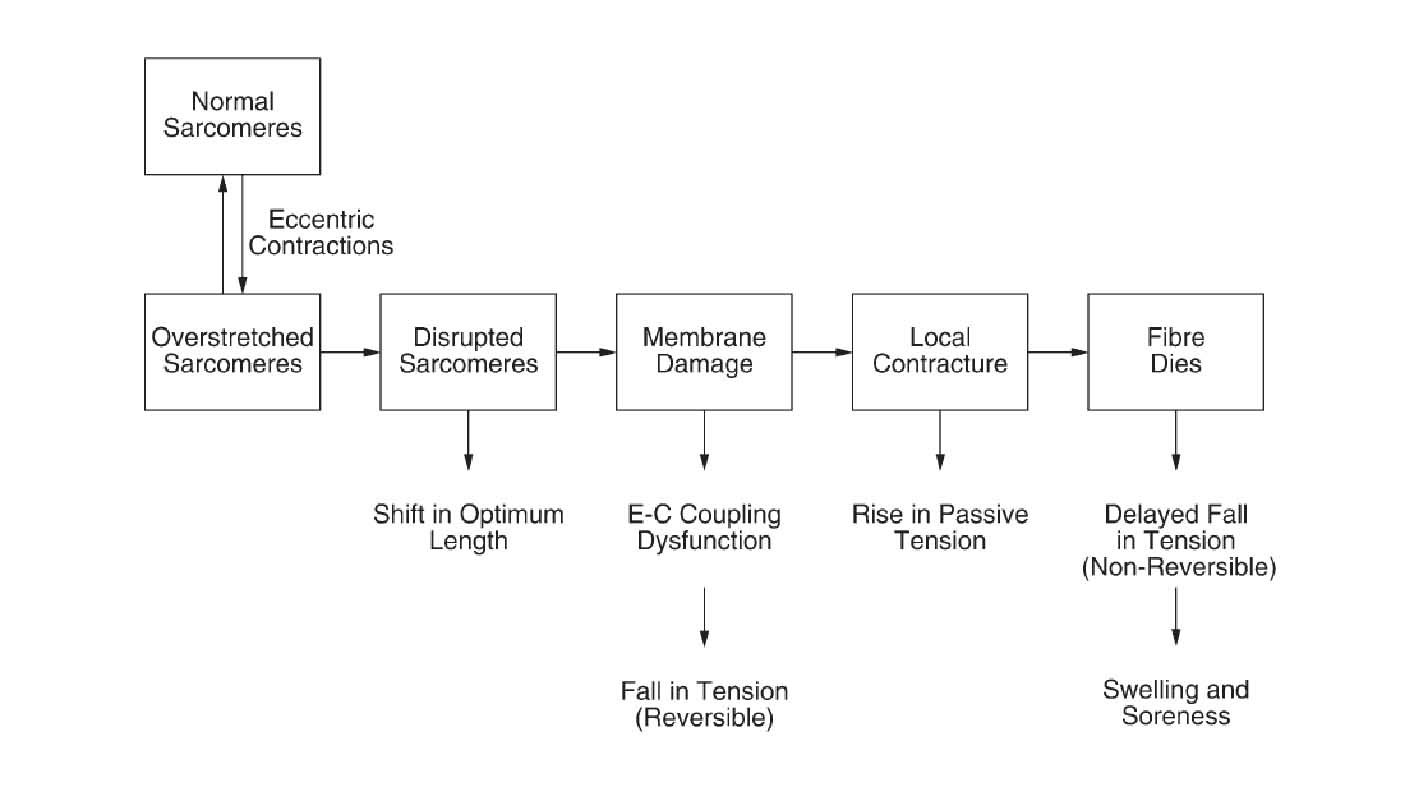
Рис.1. Постулируемый ряд событий, приводящих к повреждению мышцы при выполнении эксцентрического упражнения
В течение активного удлинения, длинные, более слабые саркомеры растянуты на спускающуюся ветвь кривой, где они удлиняются быстро, неудержимо, пока они не вне наложения миофиламентов и напряжение в пассивных структурах не остановила далее удлинение. Повторное сверхнапряжение саркомеров приводит к их разрушению. Мышечные волокна с разрушенным саркомерами, расположенными последовательно со все еще функционирующим саркомерами показывают изменение в оптимальной длине характеристики длина-напряжение в направлении более длинных длин мышцы. Когда область разрушения является достаточно большой, это приводит к мембранному повреждению. Это могло быть предусмотрено как двухэтапный процесс, начинаясь с разрывания t-трубочек. Любое падение напряжения в этом месте может быть обратимо посредством кофеина (см. текст). Это сопровождалось бы повреждением саркоплазматического ретикулума, выходу ионов Ca2+ и вызову местной контрактуры раны. Это, в свою очередь, подняло бы пассивное напряжение мышцы. Если бы повреждение было достаточно обширно, то части волокна, или целого волокна, погибли бы. Это падение в напряжении не было бы восстанавлено кофеином. Продукты деструкции мертвых и умирающих ячеек привели бы к местному воспалительному ответу, связанному с отеком ткани и болевыми ощущениями.
Концептуально более трудно предусмотреть повреждение процесса сцепления E-C как первичный случай. Однако, наблюдения Takekura et al. (2001) над неправильными системами t- трубочек после эксцентрического упражнения могли бы обеспечить подтверждение этой гипотезы. В этом случае начало повреждения связано с разрывом t-трубочки. Порванные концы трубочки привели бы к инактивации некоторых саркомеров. Если бы такие саркомеры были сконцентрированы в специфических миофибриллах, то это могло бы привести к падению напряжения. Это напряжение восстанавливалось бы кофеином, но не потенциацией. Если бы инактивированные саркомеры были бы рассеяны наугад среди миофибрилл, ситуация была бы подобной тому, что происходит при первичном разрушении саркомера. Таким образом, ультраструктурное, неоднородное распределение длины саркомеров было бы соблюдено и механически. Изменение в оптимальной длине для активного напряжения произошло бы в направлении больших значений длины мышцы. Главная трудность в применении этой гипотезы заключается в том, что трудно объяснить, почему t-трубочки должны быть первичным участком повреждения и почему это происходит только при длинах саркомера, находящихся вне оптимума. Обратная последовательность, при которой все начинается с разрушения саркомера и затем, приводит к повреждению t- трубочки также согласовывается с наблюдениями Takekura et al. (2001).
СТРУКТУРНЫЕ ПРИЗНАКИ
В настоящее время не подлежит сомнению, что эксцентрическое упражнение приводит к структурным повреждением мышцы (Friden et al. 1981; Newham et al. 1983). Большинство свидетельств этого повреждения получено посредством оптического микроскопа. Они показывают, что при повреждении происходит: нарушение порядка «регистрации саркомеров», «вытекания» Z-линии, появление областей перенапряжения саркомеров или полусаркомеров, региональную дезорганизацию системы t- трубочек и миофиламентов (см.обзор Morgan & Allen (1999).
Точные детали процесса разрушения саркомеров после эксцентрических сокращений остаются объектом исследования и новых гипотез. Одним из объектов исследования является упругая нить белка титина, который «ставит на якорь» толстые нити к дискам Z, или структурный белок десмин, который связывает смежные Z-диски (Аллен, 2001). Существуют данные, что из-за маленьких ошибок при размещении, толстые и тонкие нити сверхрастянутых саркомеров могут действовать (бодать) друг против друга. Инактивация некоторых саркомеров от места повреждения до системы t-трубочек может также играть роль. Безотносительно точных деталей, есть свидетельство перенапрягшихся саркомеров и полу-саркомеров в мышце, которая подвергается эксцентрическим сокращениям (см., например, Brown & Hill, 1991; Lieber et al. 1991; Wood et al. 1993; Talbot & Morgan, 1996). Недавно было показано на долях намагниченных (permeabilised) мышечных волокон крысы, что области, где были расположены длинные саркомеры до активного растяжения содержали большинство разрушенных саркомеров после растяжения. Эти разрушенные саркомеры были более длинными чем остальные (Macpherson et al. 1996).
Диагностическая структурная особенность разрушения после эксцентрического упражнения – присутствие сверхрастянутых полу-саркомеров, со смежным полусаркомерами, сильно сокращенными (для примера, см. Brown & Hill, 1991; Talbot & Morgan, 1996; Macpherson et al. 1997). Эта последовательная особенность побудила нас размышлять о расположении упругих нитей внутри саркомеров. Титин, как полагают, ставит на якорь концы толстых нитей к Z-диску (Horowits, 1999). В простой механической модели саркомера, если бы титин был приложен только к концам толстых нитей, два полу-саркомера вели бы себя независимо друг от друга, и сверхрастяжение одной половины не должно было бы привести к сокращению другой половины (рис. 2). Если, однако, второй упругий элемент охватывает всю длину саркомера, мы увидим те структурные изменения, которые видны на электронном микроскопе. Таким образом, сокращенный полусаркомер вызывает растягивание сверхрастянутого полусаркомера (рис. 2).

Рис.2. Постулируемое распределение упругих нитей в саркомерах. Две верхние диаграммы рассматривают саркомер с упругими нитями, привязывающими только концы толстых нитей к Z-линиям. Числа указывают распределение напряжения. Полное напряжение установлено в 80 % максимума указывает, что саркомер находится на спускающейся ветви зависимости длина-напряжение. Когда один полусаркомер становится сверхрастянутым, в результате эксцентрического сокращения, напряжение, которое переносят его миофиламенты, падает до нуля и полное напряжения переносит упругая нить. Другой полусаркомер незатронут, так как его изометрическая способность напряжения остается той же самой. Когда второй упругий элемент включен в модель, тот, который охватывает полную длину саркомера (3 диаграммы, расположенные внизу), сверхрастяжение одного полусаркомера принуждает 50 % напряжения быть распределенными упругому элементу последовательно с толстыми нитями, в то время как, из-за его пропорционально меньшего расширения, тот охват саркомер имеет меньше (30 %). Это также внесет 30 % напряжения к другому полусаркомеру, остающиеся 50 %, распределяемых между рядом упругий элемент (5 %) и поперечные мостиков (45 %). Так как изометрическая способность развивать напряжение этих полусаркомеров остается в 70 %, это сокращается, пока напряжение в элементе ряда не упала к нолю, и наложение миофиламентов находится где-нибудь на возрастающей ветви, производя 60 % напряжения, оставляя 20 % в упругом элементе, охватывающем саркомер. Это то, что видно под электронным микроскопом, один полусаркомер, сверхрастянутый вне наложения, другой наполовину очень короткий. Это модель указывает, что пассивное напряжение в целом саркомере становится существенным, когда одна половина саркомера становится сверхрастянутой.
Еще меньше доступной информации, о структурном повреждении системы сцепления E-C. В недавнем исследовании крыс, которые выполняли бег по наклонной плоскости, в мышцах передних лап было обнаружено множество ультраструктурных отклонений, включая больше растянутых частей системы t-трубочек, изменения в расположении триад, группы caveolar и смещение частей системы t-трубочек с относительно терминальных цистерн (Takekura et al. 2001). 2001). По мышечному волокну были «разбросаны» в беспорядке мембранные системы. Эти данные соответствовали гипотезе, о центральном повреждении саркомера, скольжению миофибрилл друг относительно друга и возможного повреждения системы t-трубочек.
СДВИГ ОПТИМУМА ДЛИНЫ САРКОМЕРА
Существует ли какое-нибудь свидетельство механических свойств мышцы, поддерживающее существование сверхрастянутых саркомеров в поврежденный мышце? Было выдвинуто предположение, что присутствие сверхрастянутых саркомеров увеличивает последовательную податливость, приводя к изменению зависимости «длина-напряжение» активной мышцы в направлении больших значений длины мышцы (Morgan, 1990). Такое изменение было сначала описано Katz (1939) а затем показано для изолированных мышечных волокон лягушки (Morgan et al. 1996), целой мышцы земноводных (Wood et al. 1993; Talbot & Morgan, 1996) и мышцы человека (Jones et al. 1997; Brockett et al. 2001). Моделируемый пример показан на рис. 3A.
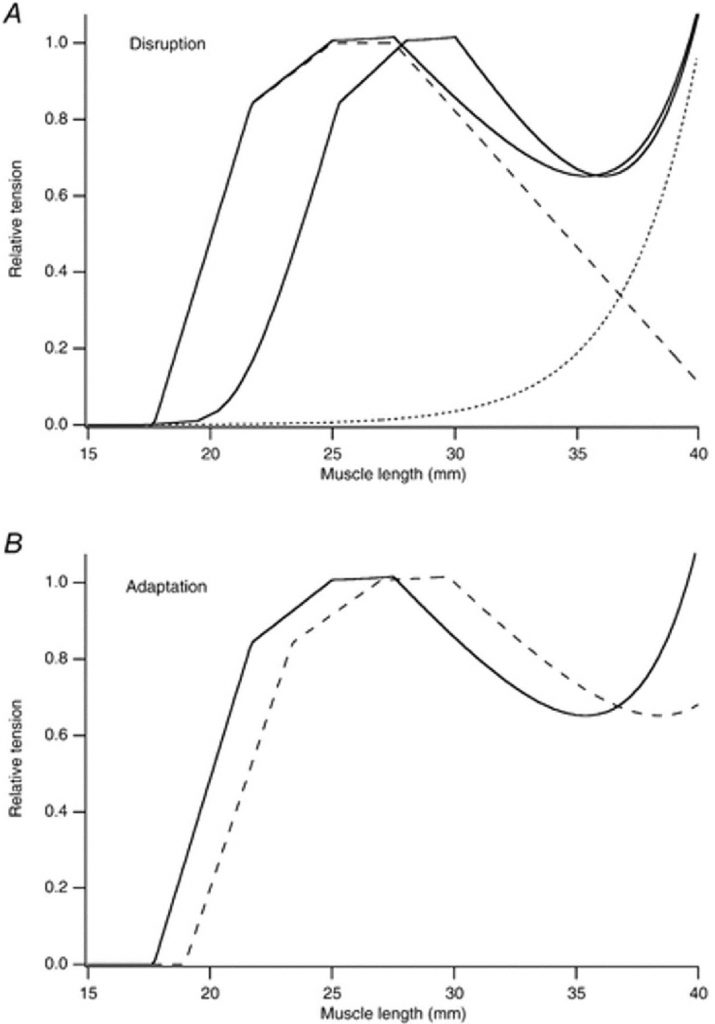
Рис.3. Изменения в механических свойствах мышцы после ряда эксцентрических сокращений.
A, разрушение саркомеров. Компьютер моделировал зависимость длина-напряжение; для саркомера. Штриховая линия; зависимость длина-напряжение для саркомера, взятое из статьи Gordon et al. (1966).
Пунктир — показательная кривая, представляющая пассивную напряженность; непрерывная линия — полная напряжение. Напряжение нормализовано относительно максимума активного напряжения. Волокно состоит из 10 000 саркомеров, имеющих среднюю длину 2,5 мкм. Исходная зависимость непрерывная кривая слева. После ряда эксцентрических сокращений 10 % саркомеров не могут развивать силу, поэтому чтобы моделировать разрушение, происходит изменение в оптимальной длине зависимости длина-напряжение на 3 мм вправо (непрерывная кривая справа).
B, адаптация мышечного волокна после повреждения, вызванного эксцентрическим упражнением. Непрерывная кривая — кривая напряжения общего количества контроля как в верхней группе, расплющенной кривой, что после того, как число саркомеров последовательно было увеличено на 10 %, не изменяя длину сухожилия. Это привело к увеличению оптимальной длины на 2 мм.
Если основной причиной дефицита напряжения после эксцентрических сокращений был отказ в сцепления E-C (Warren et al. 1993), приводя к сниженному, но однородному выходу ионов Ca 2+ , то изменение в отношении «длина-напряжение» могло бы попасть на сниженный уровень активации, так, чтобы мышцу нужно было бы больше растянуть, чтобы она достигла максимальной активации (Endo, 1973). Существуют доказательства, что такое объяснение не всегда правильно. Для изолированного мышечного волокна лягушки (Morgan et al. 1996) и целой мышцы крысы (Рис. 4), могут быть найдены примеры, где кривые «длина-напряжение» до и после эксцентрического сокращения пересекаются при значительной длине (Katz, 1939; see also Brockett et al. 2001a, b).
При этих больших значениях длины напряжение после выполнения упражнения выше, чем до упражнения. Таким образом, чтобы объяснить изменение в кривой (рис. 4) нельзя использовать ссылку на неполную активацию мышцы.
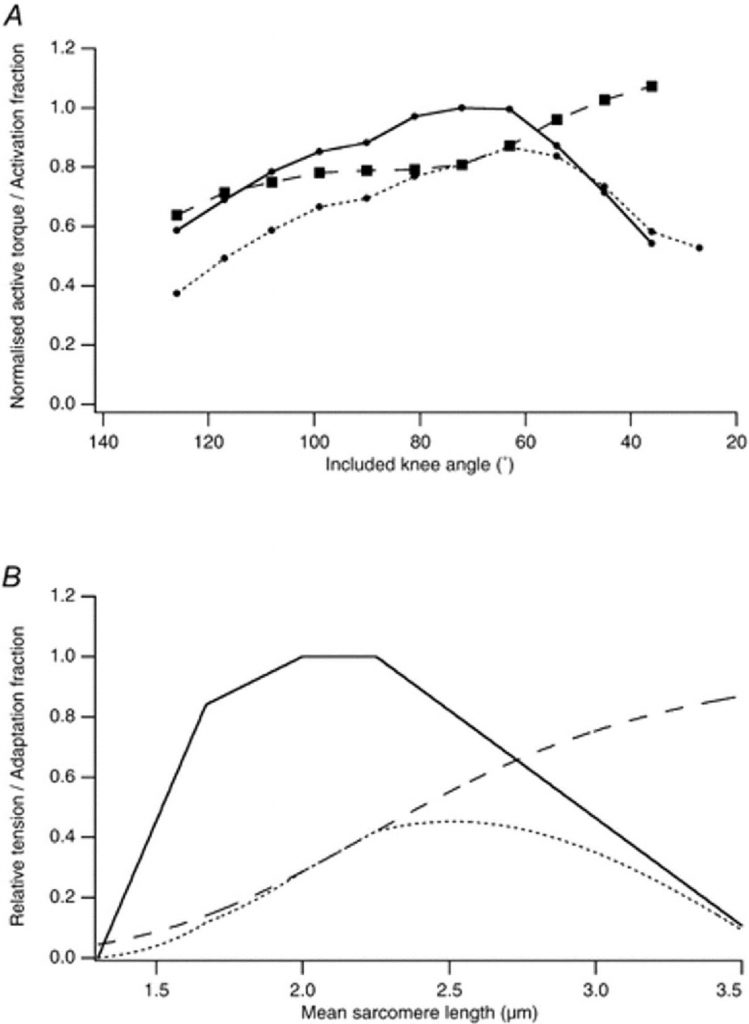 Рис.4. Изменения в механических свойствах мышцы после ряда эксцентрических сокращений. А, разрушение саркомеров. Компьютер моделировал зависимость длина-напряжение для саркомера. Штриховая линия — зависимость длина-напряжение для саркомера, взятое из статьи Gordon et al. (1966). Пунктир — показательная кривая, представляющая пассивную напряженность; непрерывная линия — полная напряжение. Напряжение нормализовано относительно максимума активного напряжения. Волокно состоит из 10 000 саркомеров, имеющих среднюю длину 2,5 мкм. Исходная зависимость -непрерывная кривая слева. После ряда эксцентрических сокращений 10 % саркомеров не могут развивать силу, поэтому чтобы моделировать разрушение, происходит изменение в оптимальной длине зависимости длина-напряжение на 3 мм вправо (непрерывная кривая справа). B, адаптация мышечного волокна после повреждения, вызванного эксцентрического упражнения. Непрерывная кривая — кривая напряжения общего количества контроля как в верхней группе, расплющенной кривой, что после того, как число саркомеров последовательно было увеличено на 10 %, не изменяя длину сухожилия. Это привело к увеличению оптимальной длины на 2 мм.
Рис.4. Изменения в механических свойствах мышцы после ряда эксцентрических сокращений. А, разрушение саркомеров. Компьютер моделировал зависимость длина-напряжение для саркомера. Штриховая линия — зависимость длина-напряжение для саркомера, взятое из статьи Gordon et al. (1966). Пунктир — показательная кривая, представляющая пассивную напряженность; непрерывная линия — полная напряжение. Напряжение нормализовано относительно максимума активного напряжения. Волокно состоит из 10 000 саркомеров, имеющих среднюю длину 2,5 мкм. Исходная зависимость -непрерывная кривая слева. После ряда эксцентрических сокращений 10 % саркомеров не могут развивать силу, поэтому чтобы моделировать разрушение, происходит изменение в оптимальной длине зависимости длина-напряжение на 3 мм вправо (непрерывная кривая справа). B, адаптация мышечного волокна после повреждения, вызванного эксцентрического упражнения. Непрерывная кривая — кривая напряжения общего количества контроля как в верхней группе, расплющенной кривой, что после того, как число саркомеров последовательно было увеличено на 10 %, не изменяя длину сухожилия. Это привело к увеличению оптимальной длины на 2 мм.
Гипотеза неоднородности саркомера предсказывает, что повреждение будет встречаться только в том случае, если длина саркомеров после растяжения будет больше оптимальной длины. Если разрушение саркомера и повреждение особенно видны на спускающейся ветви кривой «длина-напряжение» мышцы, следовательно индикаторы повреждения зависят от длины мышцы. Это действительно имеет место. И для мышцы крысы и для жабы сдвиг оптимальной длины и падение активного напряжения зависят от степени растяжения мышцы (Lynn & Morgan, 1998; Talbot & Morgan, 1998).
Изменение оптимальной длины при активном напряжении, в направлении больших длин мышцы, является довольно косвенным индикатором увеличенного сопряжения мышцы, после выполнения упражнения. Есть ли какие-либо другие механические изменения в мышце, совместимые с увеличением сопряжения? В оригинальном обзоре Katz (1939) показал в 2-3 раза более медленное повышение изометрического тетанического напряжения и уменьшение соотношения twitch:tetanus, совместимого с «частичным преобразованием активных сокращений в пассивную упругую ткань». В недавней серии экспериментов на медиальной головке икроножной мышцы анестезированной кошки было найдено, что, после ряда эксцентрических сокращений повышение напряжения в ответ на растяжение пассивной мышцы было отсрочено, по сравнению с реакцией перед сокращениями (Whitehead et al. 2001). Наша интерпретация этого факта состоит в том, что эксцентрические сокращения привели к сверхрастяжению и разрушению некоторых саркомеров. Когда мышца расслабилась, некоторые перенапрягавшие саркомеры не пересчитали (Talbot & Morgan, 1996), и это означало, что соседние саркомеры возвратились к более короткой длине, чем до сокращения. Мышца должна была бы поэтому быть растянута больше до того, как пассивное напряжение в этих более коротких саркомерах повысится до измеримых уровней.
СНИЖЕНИЕ НАПРЯЖЕНИЯ
Важный индикатор повреждения после периода эксцентрического упражнения — понижение активного напряжения. Оно может достигать 60 %, но нужно помнить, что снижение напряжения включает компонент метаболического утомления, которое применимо ко всем формам упражнения, а не только эксцентрического упражнения. Предполагается, что любая оценка снижения в напряжении при эксцентрическом упражнении должна использовать в качестве контроля сопоставимый период концентрического упражнения, но нужно принять во внимание различие в энергетической стоимости концентрических и эксцентрических сокращений (Allen, 2001).
Разрушение саркомеров, которые лежат последовательно с функционирующими саркомерами, не приведет к падению напряжения, если длина волокна была приспособлена к новой, более длинной оптимальной длине. Но если повреждение будет распространяться поперек миофибриллы к другим миофибриллам, в этот процесс будут вовлекаться мембранные структуры, что приведет к повреждению и ухудшению процесса сцепления E-C. Любое снижение напряжения в этой точке может быть восстановлено контрактурой кофеина. Впоследствии была бы достигнута точка, где часть волокна, или все волокно погибло. С этим согласуется сообщение о вторичном, отсроченном падении напряжения (Jones et al. 1989; Faulkner et al. 1993; MacIntyre et al. 1995). У человека было обнаружено, что начальное падение напряжения после эксцентрических упражнений сопровождается медленным повышением его через 2-4 часа, что, по-видимому, характеризует восстановление после метаболического истощения. Однако после 24 часа наблюдается вторичное падение напряжения (MacIntyre et al. 1995). Возможно, это связано с тем, что некоторые волокна погибают, и поэтому этот компонент напряжения не будет восстанавливаем посредством кофеина (рис. 1).
Делая измерения снижения напряжения после эксцентрического упражнения, не только важно рассмотреть любые эффекты метаболического утомления, но и последствия изменения в оптимальной длине активного напряжения. Оптимальная длина перед упражнением будет короче, чем после упражнения из-за сдвига и это может привести к недооценке остаточного напряжения. Это, вероятно, будет, также способствовать наблюдаемому изменению в зависимости «частота-сила» (Newham и др. 1983).
РАЗЛИЧНЫЕ ТИПЫ МЫШЕЧНЫХ ВОЛОКОН
Во многих исследованиях предполагается различная восприимчивость мышц и их частей к повреждению после эксцентрических сокращений. Вопрос, который поднимается неоднократно, касается уязвимости различных типов ДЕ в мышцах смешанного состава, состоящих из медленных и быстрых мышечных волокон. Предполагалось, что при локомоции повреждаются медленные мышечные волокна (Armstrong et al. 1983; Mair et al. 1992). Это связано с низким порогом активации и выполнением важных задач поддержания позы. Когда мышцы смешанного, медленно-быстрого состава волокна были подвергнуты максимальному активному растяжению, большие, быстрые ДЕ были более уязвимыми, и было выдвинуто предложение, что это происходит из-за их низкой окислительной способности (Friden & Lieber, 1998), или более высокого напряжения, которые они генерируют (Appell и др. 1992). В другом недавнем исследовании, демонстрирующем преимущественное повреждение быстрых оксидативно-гликолитических волокон предполагалось, что, за это отвечает фенотип волокна или более низкая способность к сокращению (Vijayan и др. 2001). Другие предложили комбинацию факторов, вовлекающих и активные и пассивные свойства мышечных волокон (Macpherson et al. 1996). В недавнем обзоре Lieber & Friden (1999) предположили, что большее повреждение быстрых волокон после эксцентрического упражнения является результатом «увеличенного напряжения и повреждения из-за небольшой длины волокна».
Важная особенность гипотезы неоднородности саркомеров — зависимость повреждения от длины мышцы, на которой выполняются эксцентрические сокращения. Это поднимает вопрос о том, что в мышце смешанного состава оптимальная длина для различных типов волокон, возможно, не одинакова, поэтому растяжение всей мышцы принудило бы некоторые волокна быть растянутыми больше на опускающейся ветви кривой «длина-напряжение» чем другие. В недавнем исследовании нашей лаборатории (J. Talbot, M. Homewood & D.L.Morgan, неопубликованные данные), быстро сокращающаяся мышца tibialis anterior и медленная мышца soleus крысы были подвергнуты ряду эксцентрических сокращений. Активные растяжения мышцы должны были «покрыть» одну и ту же часть кривой «длина-напряжение» каждой мышцы. Изменения в оптимальной длине для активного напряжения наблюдались в обеих мышцах, указывая, что сокращения произвели повреждение в обоих типах волокна. Размер изменения не существенно отличался между этими двумя мышцами. В результате было выдвинуто предположение, что состав мышечных волокон разного типа, не является определяющим фактором степени повреждения при эксцентрических сокращениях, если сокращения покрывают эквивалентный диапазон длины мышцы.
Однако, все еще оставались необъясненными сообщения других авторов относительно восприимчивости к повреждению различных типов волокон в пределах одной и той же самой мышцы, имеющей смешанный состава волокон. Во втором эксперименте (Brockett и др. 2001b), изучались быстрые и медленные ДЕ медиальной головки икроножной мышцы кошки. Было найдено, что у большинства быстрых ДЕ единиц оптимальная длина, при которой развивается максимальное напряжение меньше, чем для целой мышцы. Наоборот, у медленных ДЕ оптимальная длина, в среднем, больше, чем оптимальная длина целой мышцы. Когда ДЕ были подвергнуты ряду эксцентрических сокращений, используя сокращения, которые начались в оптимуме целой мышцы, все единицы показали изменение зависимости «длина-напряжение», что свидетельствовало о повреждении. Однако, медленные ДЕ показали меньшее изменение чем быстрые ДЕ. Статистический анализ показал, что оптимальная длина ДЕ, при которой развивается максимальное напряжение, относительно оптимума целой мышцы, была лучшим индикатором, чем тип ДЕ, отражающим восприимчивость ДЕ к повреждению от эксцентрического упражнения. Различие в оптимальной длине двух типов ДЕ, как предполагалось, происходило из-за различий в числе саркомеров последовательно расположенных в мышечном волокне. Это, в свою очередь, как думали, касалось роли различных типов ДЕ в gastrocnemius во время позы и ходьбы у кошки (Brockett и др. 2001b). Полученные результаты теперь должны быть подтверждены экспериментами на других мышцах смешанного состава.
ПОВЫШЕНИЕ ПАССИВНОГО НАПРЯЖЕНИЯ
Как известно в течение некоторого времени после п nbsp; медиальной головки икроножной мышцы кошки. Было найдено, что у большинства быстрых ДЕ единиц оптимальная длина, при которой развивается максимальное напряжение меньше, чем для целой мышцы. Наоборот, уериода эксцентрического упражнения в мышце повышается пассивное напряжение. Для флексоров локтя человека на это указывает то, что расслабленная рука, принимает немного согнутое положение (Jones et al. 1987). Когда была измерена жесткость мышцы, было установлено, что после упражнения она была в два раза выше и оставалась такой повышенной в течение последующих 4 дней (Howell et al. 1993). Непосредственное повышение жесткости после упражнения, как постулировалось, следовало из активизированного растяжением выхода ионов кальция (Ca 2+ ) (Howell et al. 1993). Другие объяснения были основаны на укорочении параллельных несократительных элементов в мышце (Howell et al. 1985; Jones et al. 1987) В схеме, показанной на рис. 1, есть пассивное напряжение, повышающееся с развитием местной контрактуры в сегментах волокна, после повышения в саркоплазме концентрации ионов кальция (Ca 2+ ), в результате повреждения мембраны.
Несколько исследований продемонстрировали увеличение во время отдыха уровня Ca 2+ в мышечных волокнах, поврежденных эксцентричными сокращениями, (Balnave & Allen, 1995; Balnave и др. 1997; Ingalls и др. 1998). Однако, в рамках разрешения метода, измерения показали, что повышенные уровни Ca 2+ были однородно распределены среди мышечных волокон (Balnave и др. 1997). Возможно, повышение Ca 2+ достаточно, чтобы вызвать низкий уровень активации, чтобы увеличить пассивное напряжение, хотя этому нет никакого прямого доказательства (Whitehead et al. 2001, p. 602).
Когда измерения проводятся по всему физиологическому диапазону, повышение пассивного напряжения после того, как ряд эксцентрических сокращений достигает максимума в длине близко к оптимуму для активного напряжения (Whitehead et al. 2001). Измерения поглощения энергии пассивной мышцей в ответ на большое, медленное удлинение и укорочение движений показали значительное повышение после эксцентрических сокращений. Предполагалось, что увеличение поглощения энергии было результатом активной циркуляции поперечных мостиков в поврежденных сегментах мышечных волокон (Whitehead et al. 2001). Есть некоторый структурный признак сокращенных сегментов волокна в мышце, поврежденной эксцентричным упражнением. Было найдено, что после бега по наклонной плоскости, волокна мышцы soleus крыс показали «растворение» Z-линии, разрушение A-диска и свертывание волокна (Ogilvie и др. 1988). Так же Friden & Lieber (1998) нашли, что в мышце кролика после эксцентрических сокращений был разрушен цитоскелет и появились области гиперсокращения.
НАБУХАНИЕ МЫШЦЫ И БОЛЕЗНЕННОСТЬ
Эксцентрическое упражнение сопровождается ощущениями неподвижности и болезненности на следующий день (Hough, 1902). Текущее представление механизма состоит в том, что повреждение на уровне саркомеров ведет, во время повторных сокращений, к более значительному ущербу и, в конечном счете, к смерти некоторых мышечных волокон. Повреждение вызывает местную воспалительную реакцию, которая сопровождается некоторым отеком. Продукты распада травмированных тканей делают чувствительными болевые рецепторы (Smith, 1991; MacIntyre et al. 1995).
Предполагал ось, что жесткость после эксцентрического упражнения происходит из-за набухания, сопровождающего повреждение. Таким образом Howell et al. (1985) предположил, что отсроченное увеличение жесткости в мышцах флексора локтя после упражнения является результатом изменений объема перимизия и эпимизия соединительной ткани. Количественная биомеханическая модель поддерживала это представление (Purslow, 1989). Однако, пассивное напряжение и изменение жесткости появляются немедленно после упражнения (Howell et al. 1993; Chleboun et al. 1998; Whitehead et al. 2001), когда еще нет никакого признака набухания. В наших экспериментах было выявлено повышение пассивного напряжения в мышцах разгибателях стопы, которое достигло своего пика сразу после упражнения. Через 24 часа после упражнения, когда достигло максимума разбухание мышцы, не было никакого значительного увеличения пассивного напряжения. Таким образом, наши эксперименты свидетельствуют о том, что нет связи между повышением пассивного напряжения и набуханием мышцы. Набухание начало стихать через 4 дня после выполнения упражнений.
Болезненность начинается приблизительно через 6-8 часов после упражнения и достигает пика приблизительно через 48 часов (MacIntyre et al. 1995; Jones et al. 1997). Настоящее представление этого механизма состоит в том, что продукты распада ткани делают чувствительным болевые рецепторы так, чтобы они ответили на стимулы, которые обычно невредны. Таким образом, мышца имеет тенденцию к местной пальпации, растяжению и сокращению. Недавно было выдвинуто предположение, что компонент DOMS от эксцентрического упражнения вовлекает большие механорецепторы, расположенные в мышечных волокнах (Barlas и др. 2000; Weerakkody и др. 2001). Было установлено, что от механорецепторов, включая мышечные веретена, начинается болевой путь к спинному мозгу.
АДАПТАЦИЯ
Мы все знаем, что результатом адаптации мышцы является то, что неподвижность и болезненность после периода упражнения становятся намного меньше, когда упражнение повторяется неделю спустя. Это известно давно (Hough, 1902; Friden et al. 1983; Schwane & Armstrong, 1983; Clarkson & Tremblay, 1988). Однако до сих пор вызывает споры механизм этого процесса (см. обзор McHugh et al. 1999).
Первоначально Morgan (1990) предположил, что при повреждении в результате эксцентрических упражнений процесс адаптации увеличивал количество саркомеров последовательно расположенных в мышечных волокнах. Как следствие, средняя длина саркомера становилась короче (рис. 3B). Поэтому только небольшая часть рабочего диапазона мышцы входила в область неустойчивости, характерную для кривой «длина-напряжение». Эта гипотеза была поддержана экспериментами, в которых крысы бегали на тредмилле по наклонной плоскости вверх и вниз (Lynn & Morgan, 1994; see also Lynn et al. 1998). Затем волокна мышцы, которая работала в эксцентрическом режиме, (vastus intermedius), были фиксированы и было подсчитано количество саркомеров после недели упражнений. Было найдено, что среднее число саркомеров в мышце, выполняющей эксцентрические сокращения было на 11% больше, по сравнению с мышцей работающей в концентрическом режиме.
Это предположение было оспорено (Koh & Herzog, 1998; см. также Koh & Brooks, 2001). Однако, противоположные результаты исследования трудно оценить, так как в исследовании мышцы кролика не было дано сведений о диапазоне изменения длины мышцы, которая растягивалась относительно оптимальной длины. Кроме того не было заявлено, сопровождался ли эксцентрический протокол сокращения какими-либо поддающимися обнаружен Существует гипотеза процесса разрушения саркомера (рис. 1). Как известно, существует область, где напряжение саркомера падает (из-за особенностей зависимости ию признаками разрушения или повреждения.
Скептицизм по механизму адаптации, вовлекающему добавление дополнительных саркомеров к мышечным волокнам, сосредоточивается относительно времени адаптации (McHugh et al. 1999). Если мышечные волокна должны быть реконструированными после повреждения эксцентрического упражнения этот процесс должен быть достаточно быстрым, чтобы быть полностью завершенным к концу недели после повреждения. В состоянии ли мышца перенести такой быстрый процесс адаптации на уровне саркомеров? Однако еще в 1973 году было показано, что иммобилизация мышцы в удлиненном положении посредством гипсовой повязки привела к увеличению количества саркомеров в мышечных волокнах в течение 5 дней (Williams & Goldspink, 1973). Это увеличение было быстро обратимо. В то время как мы еще не понимаем точные детали, на клеточном уровне, событий, которые приводят к адаптации количества саркомеров (Goldspink, 1998; см. также Wretman и др. 2001), скорость процесса очевидно достаточна, чтобы составлять адаптацию, наблюдаемую после эксцентрического упражнения.
Если мышца приспосабливается к повреждению от эксцентрического упражнения посредством увеличения количества саркомеров в мышечных волокнах, какие применения это имеет для мышечно-сухожильных отношений? Мы попытались смоделировать эту ситуацию (рис. 5). Считайте мышцу составленной из мышечных волокон, содержащих 10 000 саркомеров и 20 мм сухожилия. В этой модели мышцы напряжение начнет увеличиваться при растяжении мышцы на 35 мм, при этом длина саркомера составляет 1,5 мкм. Напряжение достигнет максимума при растяжении 45 мм или длине саркомера в 2,5 мкм. Если цель состоит в том, чтобы сместить кривую «длина-напряжение» мышцы в направлении больших значений длины, чтобы обеспечить защиту против повреждения от эксцентрического упражнения, простая адаптация должна будет увеличить длину сухожилия на 5 мм. Это сместило бы оптимальную длину на 5 мм. Однако, напряжение было бы менее хорошо поддержано на короткой длине мышцы, падая до нуля в 40 мм, а не в 35 мм. Если бы число саркомеров в волокне было увеличено на 20 %, не изменяя длину сухожилия, то необходимое изменение в оптимальной длине было бы все еще получено, но рабочий диапазон мышцы был бы уменьшен на 3 мм. Только если число саркомеров будет увеличиваться и в дальнейшем, могло быть достигнуто сокращение сухожилия и изменение в оптимальной длине и обслуживание оригинального рабочего диапазона мышцы (рис. 5). Практически, маловероятно, что сухожилие в состоянии быть реконструированным в течение недели, хотя это остается возможностью для адаптации в дальнейшей перспективе.
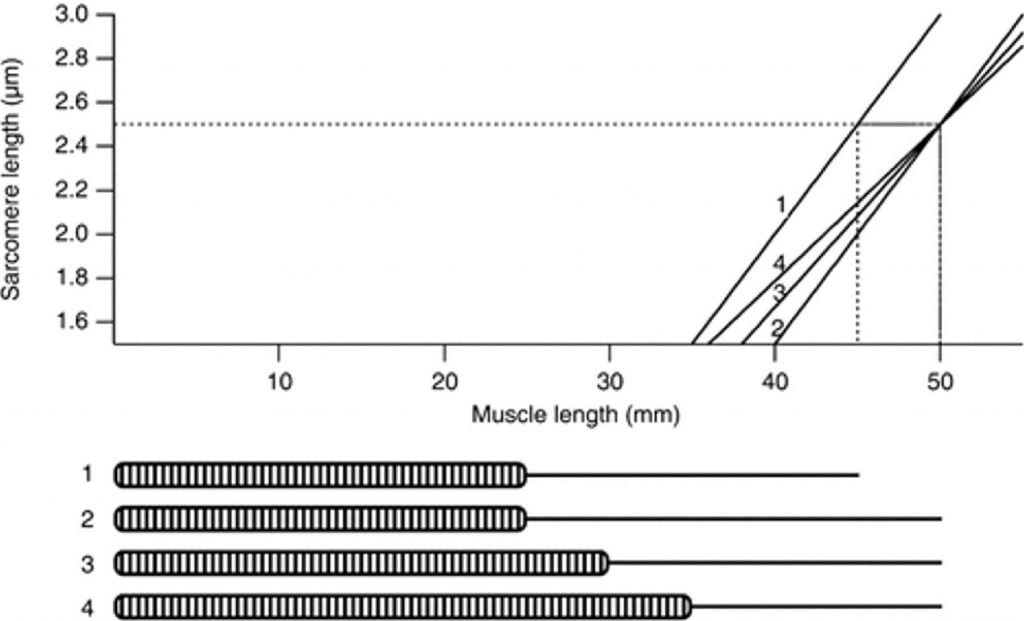 Рис.5. Кривая длина-напряжение для саркомера
Рис.5. Кривая длина-напряжение для саркомера
Отношение между длиной саркомера и длиной мышцы для теоретических мышечных волокон, включающих различное количество саркомеров и различную длину сухожилия. Предполагается, что оптимальная длина саркомера — 2.5 мкм. Для мышечного волокна, имеющего 10 000 саркомеров и 20 мм сухожилия (волокно № 1) напряжение начинает повышаться в 35 мм, и оптимум достигается в 45 мм (штриховая линия). Изменение в оптимальной длине для активного напряжения на 5 мм в направлении больших значений длин (штриховая линия) может быть достигнуто, увеличивая длину сухожилия до 25 мм (волокно № 2). Недостаток состоит в том, что активное напряжение не развивается, пока мышца не растянется на 40 мм, то есть, рабочий диапазон длин мышцы будет уменьшен. Увеличение числа саркомеров от 10 000 до 12 000, и уменьшение длины сухожилия д 20 мм (волокно № 3) производят необходимое увеличение оптимальной длины и приводят к меньшему сокращению рабочего диапазона мышцы, где напряжение начинает повышаться в 39 мм. Увеличение числа саркомеров далее к 14 000, в то же самое время уменьшая длину сухожилия до 15 мм (волокно № 4) приводит к самому удовлетворительному результату, необходимому изменению в оптимальной длине на 5 мм и сокращению рабочего диапазона только на 1 мм.
Есть ли какой-либо признак изменения в кривой напряжения длины как признак адаптации мышцы к повреждению от эксцентрических сокращений? Во-первых, важно различить два вида изменения, которое может встречаться при выполнении эксцентрических упражнений. После периода непривычного упражнения будет изменение в оптимальной длине в результате увеличения последовательного компонента из-за разрушенных саркомеров (рис. 3A). Это сопровождается вторым, отсроченным изменением, представляющим адаптацию, увеличивая количество саркомеров (рис. 3B). В мышцах земноводных, через 6 часов после повреждения от эксцентрических сокращений, начальное изменение в оптимальной длине полностью изменилось, и оптимум возвратился к своему значению, которое он имел до выполнения упражнения (Jones et al. 1997). Мы предполагаем, что после эксцентрических упражнений, в разрушенных саркомерах некоторых мышечных волокон в течение долгого времени будет восстанавливаться нормальная структура миофиламентов и способности произвести напряжение (Talbot & Morgan, 1996). В других волокнах области разрушения, возможно, прогрессировали до более серьезного поражения. Такие поврежденные волокна больше не будут выполнять сокращение и поэтому не могут способствовать изменению в кривой «длина-напряжение». Оба из этих факторов приведут к аннулированию изменения.
В исследовании трехглавой мышцы голени человека, было возможно идентифицировать связанный с повреждением сдвиг в оптимуме зависимости «угол-вращающий момент» в направлении больших значений длин мышцы после периода эксцентрического упражнения, но все восстановилось через два дня после упражнения (Jones et al. 1997). Никаких последующих сдвигов из-за адаптации обнаружено не было. В последующем исследовании группы мышцы задней поверхности бедра был продемонстрирован длительный сдвиг в оптимуме угла (Brockett и др. 2001a). Здесь, в то время как тренировочный эффект был ясно виден, не было возможно идентифицировать ни точку начала, ни любое аннулирование от изменения, относящегося к повреждению. По-видимому, в таких ситуациях, время восстановления после начального изменения и начало процесса адаптации могут наложиться.
Во время экспериментов на животных было показано, что у волокон мышцы vastus intermedius крыс, тренируемых бежать под гору в течение недели, было больше саркомеров, чем у группы, которая выполняла бег в гору. В конце тренировки, в ответ на острый ряд эксцентрических сокращений, начинающихся с того же самого коленного угла, группа, тренируемая в беге под гору показала меньшее изменение в оптимальном угле для вращающего момента чем обучаемая в гору группа. Меньшее изменение считали показанием относительно меньшего количества повреждения (Lynn et al. 1998).
Есть также доказательство процесса адаптации, имеющее ограничения в противоположном направлении (Whitehead et al. 1998). Группа людей должна была выполнять концентрические упражнения с трехглавой мышцей одной ноги, а другая нога выполняла роль контроля. После этого обе мышцы тренировались с использованием эксцентрических упражнений. Через неделю после эксцентрических упражнений концентрически тренируемые мышцы показали больший сдвиг оптимума, который свидетельствовал о большем повреждении, чем мышцы контроля. Предполагалось, что во время концентрического упражнения мышечные волокна могут потерять саркомеры, приводя к большей уязвимости, от повреждений от эксцентрического упражнения.
ОРГАНЫ КИНЕСТЕЗИИ
В то время как довольно много известно об эффектах утомления от упражнения при местном рефлекторном действии мышечных афферентов (см. обзор, Gandevia, 2001), вопрос о том, повреждает ли эксцентрическое упражнение мышечные рецепторы остается открытым. Поэтому обсуждение будет ограничено двумя основными мышечными рецепторам мышцы: мышечным веретеном и сухожильным органом.
В позной задаче положения локтя, выполненной после периода эксцентрического упражнения для флексоров локтя одной руки, было найдено, что тренируемая рука последовательно принимала более согнутое положение по сравнению с положением нетренируемой руки (Saxton и др. 1995). Учитывая, что главные проприорецепторы, сигнализирующие о положении конечности являются мышечными веретенами (Gandevia, 1996; Proske и др. 2000), предполагается, что сигнал от мышечных веретен увеличился в результате упражнения. Таким образом, чтобы получить такой же уровень проприоцептивного сигнала, тренируемая мышца должна была быть растянута меньше, чем мышца контроля. Текущее представление состоит в том, что уровень активности покоя от мышечных веретен сигнализирует о длине мышцы и, таким образом, о положении локтя. Если бы в результате упражнения некоторые интрафузальные волокна мышечных веретен были повреждены и развили контрактуру повреждения, это, как ожидалось бы, повысило бы уровень активности покоя для данной длины мышцы и так объяснило бы вышеупомянутые результаты.
Второй эксперимент Brockett и др. (1997) использовал скорее более умеренный режим упражнения, и флексоры локтя одной руки выполняли эксцентрические сокращения, в то время как флексоры другой руки выполняли за тот же самый период концентрические сокращения. Было получено только маленькое, переходное понижение напряжения, после упражнения, которое свидетельствовало о минимальных повреждениях мышцы, но через 48 часов у руки, выполняющей эксцентрические упражнения было более разогнутое положение по сравнению с концентрически тренируемой рукой Этот результат был противоположен данным Saxton и др. (1995). Объяснение, вероятно, касается различий в серьезности упражнений. Умеренное эксцентрическое упражнение, как можно ожидать, произведет небольшое разрушение саркомеров, приводя к увеличению их последовательного соответствия. Это, в свою очередь, увеличило бы порог длины для напряжения (Whitehead et al. 1998) и мышца должна будет быть более растянута, чтобы достигнуть уровня возбуждения мышечных веретен. Если бы мышечные волокна фактически были бы повреждены, то был бы получен противоположный результат. Эти виды суждений должны теперь быть проверены на единственных, идентифицированных рецепторах во время экспериментов на животных.
Некоторые наблюдения были также сделаны на сухожильных органах после периода эксцентрического упражнения (Gregory et al. 2001). У анастезированных кошек медиальная головка икроножной мышцы была подвергнута ряду удлиняющих сокращений и была произведена оценка реакции сухожильных органов при пассивном и активном изменениях напряжения до и после упражнений. Было найдено, что в ответ на медленное растяжение после упражнения, сухожильные органы давали залп при более короткой длине мышцы. Это было приписано повышению пассивного напряжения целой мышцы, которое наступало после упражнения. У одного животного образец, состоящий из шести сухожильных органов сигнализировал об этом увеличении. В связи с этим было предположено, что повреждение, производящее повышение пассивного напряжения, широко распространилось по мышце. Однако порог напряжения и сухожильная чувствительность органа не изменили предположение, что упражнение нарушило нормальное функционирование рецепторов.
КЛИНИЧЕСКИЕ ПРИМЕНЕНИЯ
Известно, что в течение некоторого времени после эксцентрических упражнений происходит повреждение мышц, наблюдаются слабость и их болезненность. Это поднимает вопрос, могут ли умеренные симптомы, которые все мы испытываем время от времени, привести к большему количеству основных повреждений? Конкретный случай — разрыв сухожилия мышц задней поверхности бедра (Brockett и др. 2001a). Клинические заключения предполагают, что травмы этого сухожилия встречались чаще всего в результате эксцентрических сокращений (Stanton & Purdham, 1989; Garrett, 1990; Kujala и др. 1997; Sallay и др. 1996). Возможно, что на некоторых элитных спортивных состязаниях, таких как соревнования по легкой атлетике, футбол и регби, микроповреждение от умеренного эксцентрического упражнения может, в результате требований к мышце во время соревнований, прогрессировать до большего повреждения. Если это так, то способ преодоления этой проблемы заключается в том, чтобы подвергнуть атлетов умеренной эксцентричной программе подготовки, чтобы произвести адаптацию, которая защищала бы мышцы от опасности их дальнейшего повреждения. Это суждение в настоящее время проверяется в нашей лаборатории.
Другая область, где эффект адаптации от эксцентрического упражнения может оказаться полезным, известен как идиопатическая ходьба (походка эквинуса). Чаще всего этой походкой обладают дети. У них происходит сильное подошвенное сгибание и они идут на «пальчиках», а не посредственно с пятки на носок. Такая ходьба иногда связана с церебральным параличом, но она появляется и при отсутствии неврологических знаков, то есть у нормальных детей. Текущее стандартное лечение, разработанное, чтобы опустить пятку на землю, включает инъекции ботулотоксина в трехглавую мышцу голени, чтобы ее расслабить, гипсовые повязки, наложенные на мышцу в согнутом назад положении, и вставку жестких пластин графита в обувь детей. Мы недавно исследовали возможность воздействия упражнения трехглавой мышцы голени, давая детям определенную программу эксцентрических упражнений мышц для разгибателя стопы, используя механизированную стремень. В то время как настоящая работа еще продолжается, предварительные данные предполагают, что в будущем инвазивное лечение может быть заменено режимами упражнения (D. L. Morgan, C. Blackburn & P. Percival, неопубликованные данные).
Наконец, мышечная дистрофия Дюшена является дегенеративной болезнью мышцы, связанной с нехваткой белка дистрофина, связанного с сарколеммой. Исследования показывают, что дистрофин-дефицитные мыши особенно уязвимы для повреждения от эксцентрических сокращений (Head et al. 1992; Moens et al. 1993). Это поднимает вопрос о том, что дегенеративные изменения в мышцах людей с дистрофией Дюшена, могут быть вызваны эксцентрическими сокращениями. Стратегии, направленные на уменьшение такого повреждения, состояли бы в том, чтобы избежать эксцентрических сокращений в целом или альтернативно предпринять программу очень умеренного, невредного эксцентрического упражнения, в надежде на повышение процесса адаптации в поврежденных мышцах.
ЗАКЛЮЧИТЕЛЬНЫЕ КОММЕНТАРИИ
Этот обзор сосредоточил свое внимание на начальной серии событий, приводящих к повреждению мышцы от эксцентрического упражнения (рис. 1). По нашему представлению, данные в поддержку гипотезы неоднородности саркомера появились совсем недавно, и поэтому требуется обзор ее текущего статуса. Главное, что зависимость «длина-напряжение» в состоянии отражать количество повреждения и различия в эффектах от концентрического и эксцентрического упражнения. Это также помогает объяснить несколько других поведений скелетной мышцы, которые до настоящего времени не могли быть удовлетворительно объяснены (Noble, 1992; Morgan, 1994; Morgan et al. 2000). Другой областью внимания в обзоре были различные симптомы, сопровождающие повреждение. У них, в будущем, может быть практическое применение. Так, чтобы определить степень повреждения после упражнения, вместо того, чтобы измерять дефицит напряжения, который осложняется метаболическими факторами, или изменением в зависимости «длина-напряжение», которое требует ряда измерений активного напряжения, существует простой, не травматичный индикатор – оценка повышения пассивного напряжения. Тренировочный эффект, оказанный периодом непривычного упражнения очень важен, потому что это представляет потенциальное средство защиты атлетов от травм мышцы. Это может также быть полезно для других клинических условий. Его механизм вовлекает добавление саркомеров к регенерации мышечных волокон, как показано экспериментами на животных. Такой механизм оказывает дальнейшую поддержку предположения, что основной процесс повреждения зависит от длины саркомера.
Источник